
| Català | Castellà |
Manel López
Béjar
Carlos López
Plana
Fernando López
Gatius
Departament de Sanitat
i d'Anatomia Animals, Facultat de Veterinària,
Universitat Autònoma
de Barcelona
Índex
1. Introducció
2. Segmentació
o divisió embrionària
3. Morulació
4. Blastulació
5. Implantació:
Eclosió de la zona pel·lúcida
Annex:
Notes sobre el terme "embrió"
Prefaci
La finalitat d'aquest atles és facilitar l'accés a material docent que serveixi per comprendre l'estructura, forma i fisiologia de l'embrió en la fase de preimplantació, quan encara es troba lliure en el tracte genital de la mare i és factible l'aplicació de diverses tècniques de reproducció assistida. L'embrió en fase preimplantacional és una estructura de difícil estudi, tant per qüestions tècniques com ètiques; es tracta d'una estructura microscòpica viva molt làbil que pot donar lloc a un individu complert. Aquestes característiques fan molt complicada la seva observació directa quan és l'objecte d'estudi d'una assignatura o curs sobre l'embriologia, tant bàsica com aplicada a la reproducció. L'embriologia és una ciència que es caracteritza per la riquesa iconogràfica dels seus continguts, molt important per a la comprensió dels mateixos. No obstant, hem constatat que existeix una reduïda disponibilitat de textos editats que continguin material iconogràfic. És per això que pensem que les Tecnologies de la Informació i de la Comunicació ofereixen una oportunitat important per a que els alumnes de diferents assignatures o cursos puguin accedir a un suport docent virtual que facilitarà la comprensió de les matèries relacionades amb l'embrió en fase preimplantacional.
L'obra que introduïm compta amb més de 100 imatges microscòpiques que il·lustren el desenvolupament embrionari en les diferents etapes de la fase preimplantacional, les quals els autors han anat recopilant durant més de 8 anys en diferents espècies, com conill, ratolí, cabra i vaca.
En el Grau de Veterinària de la Universitat Autònoma de Barcelona s'ofereix una assignatura optativa vinculada a las biotecnologies embrionaries, per a la qual s'ha preparat especialment aquesta obra. En aquesta assignatura coincideixen matriculats alumnes procedents de diverses titulacions, no tan sols de Veterinària, per als que pensem que potser útil tenir accés a imatges i text que facin referència a les diferents etapes del desenvolupament embrionari preimplantacional. L'objectiu dels autors és que aquesta obra pugui ser utilitzada com a forma d'ensenyança bimodal, és a dir com a complement de l'ensenyança presencial, per a la qual cosa, l'obra serà ampliada en el futur per incloure les més importants tècniques de reproducció assistida, com són la producció in vitro d'embrions i la crioconservació embrionària, sense oblidar tècniques de futur com la clonació, tant terapèutica com reproductiva.
Indicar també que l'ús d'aquesta obra pot fer-se extensiu a totes aquelles persones interessades en el desenvolupament embrionari preimplantacional, tant animal com humà degut a les grans semblances que posseeixen entre ells els embrions de diferents espècies de mamífers en aquesta fase de la vida.
Per últim, volem
agrair la valuosa i inestimable ajuda que ens ha proporcionat el nostre
amic i company Dr. Joaquim Castellà, professor de la Unitat Docent
de Parasitologia i Malalties Parasitàries de la nostra Facultat,
moltes vegades una ajuda que ha anat més enllà del caràcter
tècnic i sense la qual no hagués pogut desenvolupar-se aquest
projecte.. També agraïm a la Srta. Vanessa Cadenas, llicenciada
en Veterinària, la col·laboració en la preparació
digital de les imatges i en la il·lusió i ànims posats
durant la realització del projecte. Així mateix, volem indicar
que en la realització de l'obra han col·laborat els Srs.
Pedro Mayor y Mar Fenech, becaris del nostre Departament, l'ajuda dels
quals volem agrair expressament.
1. Introducció
"There are simple as well as complex ways of viewing the early development of mammals. In one perspective, the embryo possesses all the geometrical elegance and simplicity of a colonial organism akin to Pandorina or Volvox. Cell numbers during cleavage are few. Blastocyst formation results in only two cell types: trophoblast and embryoblast. The tasks required of the embryo during the preimplantation interval are seemingly straightforward: increase the copies of the genome; segregate the early descendants of the zygote into the two necessary cell types; make contact with the uterine lining. The complexities of organogenesis and the finer aspects of cell differentiation lie ahead. Simplicity and success seem to be the keynotes of this period."
Cole Manes, 1975. Genetic and Biochemical Activities in Preimplantation Embryos. In: CL Markert and J Papaconstantinou (ed). The Developmental Biology of Reproduction. Academic Press, New York.
De la cita anterior, realitzada ja fa uns anys pel Professor Manes de la Universitat de Colorado, s'extreuen dues visions ben diferenciades, però que conviuen harmònicament, del desenvolupament embrionari preimplantacional. Aquest desenvolupament pot considerar-se com simple, alhora que elegant, si atenem a l'aparença de què es basa principalment en divisions mitòtiques successives que van augmentant el nombre de cèl·lules de l'embrió. Això donaria la visió de l'elegància de formes i la simetria, en moltes etapes, que manté l'embrió durant la seva fase preimplantacional. La visió complexa d'aquest desenvolupament arriba quan considerem que a partir d'una única cèl·lula s'obté tot un ésser viu amb la seva complexitat d'aparells i sistemes, òrgans, teixits i cèl·lules. Sense dubte, la complexitat predomina sobre la simplicitat morfològica del procés, principalment perquè encara avui dia hi ha molts aspectes del desenvolupament embrionari que resten per dilucidar. Malgrat avenços importants com el coneixement de què existeixen factors citoplasmàtics de l'ovòcit que són capaços de provocar la reprogramació d'un nucli provenint d'una cèl·lula diferenciada, "oblidant tota la seva vida anterior", encara no hi ha explicacions per molts fenòmens que tenen lloc des de què un ovòcit és ovulat fins que l'embrió format s'implanta en l'úter matern.
L'embrió en
fase de preimplantació ha sigut objecte de nombrosos estudis descriptius
i experimentals, principalment degut a que és aquesta la fase on
el concepte posseeix una major independència del contacte físic
matern. La primera descripció precisa del desenvolupament embrionari
en fase de segmentació i blastulació data de 1875 i va ser
realitzada en embrions de conill per van Beneden. No obstant, són
molts els aspectes bàsics de la fisiologia, metabolisme i control
genètic de l'embrió que resten per discernir. L'avenç
en l'obtenció de coneixements de la bioquímica i fisiologia
embrionàries ha sigut més lent que en altres espècies
no mamíferes, com l'eriçó de mar o la granota, motivat
principalment per la reduïda talla i reduït nombre disponible
dels embrions de mamífers.
D'una forma general,
els principals fenòmens que tenen lloc durant la fase de preimplantació
són, en primer lloc, l'augment del nombre cel·lular, per
tant un augment de les còpies del genoma embrionari i, en segon
lloc, la diferenciació de només dos tipus cel·lulars
a través de la blastulació. A més, un últim
fenomen que té lloc en l'estadi de blastocist és el contacte
amb la mucosa uterina per a la seva implantació.
L'ovòcit dels
animals mamífers, encara què petit comparat amb els ovòcits
d'altres vertebrats, tal com peixos, amfibis, rèptils i aus, és
una de les cèl·lules més grans del cos animal. Abans de la fecundació és una de les cèl·lules
més inactives, amb una tassa metabòlica escassament més
alta que la d'un teixit poc actiu metabòlicament com és l'ossi. És cert
que aquesta cèl·lula ja posseeix individualitat, en el sentit
que el seu complement de gens constitueix una selecció única
a partir del conjunt maternal, però la seva informació genètica
no ha estat encara utilitzada, i fins aquest moment, en tots els seus caràcters
s'assembla a altres cèl·lules maternes. Tres a quatre dies
desprès de la fecundació, l'embrió pot tenir menys
d'un centenar de cèl·lules, però pateix uns canvis
importants. La tassa metabòlica és ara tan alta com la del
teixit maternal més actiu metabòlicament, com és la
retina. Els patrons de síntesi d'ARN i la capacitat de síntesi
proteica són ara similars als de les cèl·lules d'un
organisme adult i les seves cèl·lules s'aproximen ja a la
talla mitja de les cèl·lules d'un organisme adult. Els gens
del propi embrió han començat a funcionar, codificant per
proteïnes que són potencialment reconegudes com a estranyes
per la mare (McLaren, 1982). Posteriorment, la diferenciació començarà donant lloc a un teixit, el trofoectoderm, altament especialitzat per interaccionar
amb la mare, i a la massa cel·lular interna, que donarà lloc
a tot l'individu.
2. Segmentació embrionària
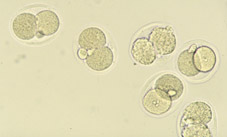
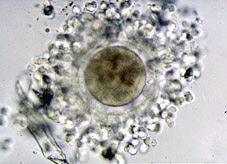
Durant el procés de fecundació té lloc el pas de l'espermatozou a través de la corona radiada i la penetració de la zona pel·lúcida de l'ovòcit (Figs 1, 2 i 3), per aconseguir-se posteriorment la fusió dels nuclis dels dos gamets (Fig 4). A partir d'aquesta fusió, totes les cèl·lules de l'organisme en desenvolupament seran portadores, normalment, de la dotació cromosòmica completa de l'espècie. L'estructura formada després d'aquest procés s'anomena zigot (Fig 5). El zigot és l'element unicel·lular a partir del qual s'origina el complex cos multicel·lular. Amb aquesta finalitat, el primer procés que esdevé és la segmentació i es caracteritza per divisions i subdivisions successives del zigot, la segmentació o divisió embrionària. L'estructura embrionària formada normalment s'anomena embrió en segmentació, o bé s'especifica el nombre de cèl·lules que el composen. Aquestes cèl·lules s'anomenen blastòmeres.
El diàmetre de l'ovòcit i del zigot és d'aproximadament 100 micròmetres en tots el mamífers (Fig 6), just el límit de visibilitat de l'ull nu. Està envoltat per una fina capa no cel·lular, la zona pel·lúcida (Fig 5). L'estadi embrionari d'una cèl·lula, degut al seu relativament gran tamany, és distingit per tenir una baixa ratio de material nuclear respecte al citoplasmàtic. Aquesta ratio, crítica pel control genètic de les cèl·lules en estadis més adults, és restablerta a un valor similar al de les cèl·lules somàtiques mitjançant el procés de segmentació, al qual moltes divisions cel·lulars successives tenen lloc sense cap increment en la massa embrionària total (Fig 7). De fet, el creixement podria ser considerat com a negatiu durant la segmentació, ja que la quantitat total de material cel·lular disminueix quasi bé en un 20% en la vaca i un 40% en la ovella, mentre que el contingut proteic de l'embrió murí decreix un 25% durant el 3 primers dies post-fecundació.
Tot el període de multiplicació cel·lular té lloc a l'interior de la zona pel·lúcida, l'embolcall acel·lular que va acompanyar a l'ovòcit en el moment de la seva ovulació. La capa de cèl·lules de la corona radiada que cobria la zona pel·lúcida de l'ovòcit no fecundat desapareix en aquest període.
La segmentació te lloc molt més lentament en mamífers que en la majoria de vertebrats inferiors o invertebrats. Els ous de granota es divideixen en aproximadament una hora, però el zigot de ratolí triga 24 hores per realitzar la seva primera divisió i de 10 a 12 hores per cada divisió ulterior. Altres embrions de mamífers mostren una similar tassa de divisió.
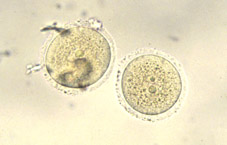
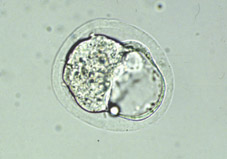
Variacions en la tassa de divisió és comú, tant entre embrions com entre les cèl·lules d'un mateix embrió. Una conseqüència és que la sincronia inicial de l'embrió en segmentació aviat es perd. Embrions amb 2 i 4 cèl·lules (Figs 8 i 9) són trobats de forma més sovint que aquells de 3 i 5 cèl·lules (Figs 10 i 11); al dia següent, l'estadi de 8 cél·lules (Fig 12, 13 i 14) predomina però la variabilitat és més gran; desprès de 4 o 5 divisions successives, poca sincronia és manté (Fig 15). La primera cèl·lula a dividir-se a partir de l'embrió de ratolí de 2 cèl·lules (Fig 16) contribueix en major manera a les cèl·lules de la massa cel·lular interna, i de forma menys marcada a la població de cèl·lules del trofoectoderm.
Durant les primeres
divisions,
cada mitosi és seguida immediatament per síntesi d'ADN en
les dos cèl·lules filles; això vol dir que la pausa
(període G1) previ a la síntesi d'ADN, la qual és
característica de les cèl·lules adultes, és
absent. Cap període G1 ha pogut ser detectat en embrions de ratolí
abans dels estadis de 8 a 16 cèl·lules (Fig
17).
3. Morulació
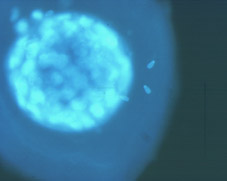
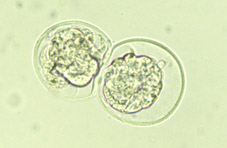
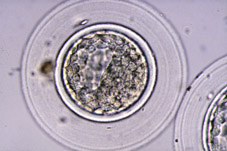
Al final del procés de segmentació, la proliferació cel·lular té com a conseqüència la formació d'un agregat de cèl·lules, íntimament unides a l'interior, que rep el nom de mòrula (Fig 18). El nombre de cèl·lules que composen la mòrula varia entre les espècies, normalment entre 16 i 64 cèl·lules (Figs 19 i 20). La mòrula ve definida pel canvi en la forma d'algunes de les seves cel·lules, ja que canvien d'esfèriques a tenir una forma de cunya, estant les cèl·lules molt pròximes entre sí per maximitzar la superfície de contacte cel·lular (Figs 21 i 22). A continuació es desenvolupen unions estretes especialitzades o complexes d'unió que aproximen encara més les cèl·lules de la mòrula, donant lloc a una pèrdua de volum total de l'embrió (Fig 23). Aquest procés globalment s'anomena compactació i dóna lloc a cèl·lules polaritzades per primera vegada, tenint diferenciats morfològica i funcionalment els pols extern i intern (Fig 21). L'estructura embrionària resultant rep el nom de mòrula compacta o compactada (Figs 24, 25, 26, 27).
Encara que existeix
una marcada variabilitat cronològica entre espècies i individus
d'una mateixa espècie, el procés de segmentació té
lloc a nivell oviductal i l'estadi de mòrula indica la fi del període
tubàric de la gestació (Figs 28
i 29).
4. Blastulació
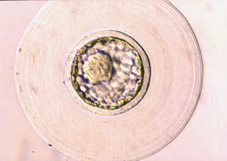
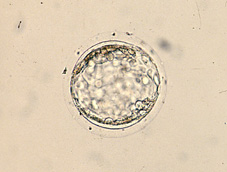

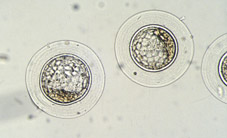
A l'inici del període uterí, les blastòmeres formadores de la mòrula compacta acumulen les seves secrecions a l'espai intercel·lular, formant unes llagunes intercel·lulars que provoquen que les cèl·lules se separin per donar lloc per coalescència a una cavitat plena de líquid que rep el nom de blastocele (Figs 30, 31, 32, 33, 34, 35, 36 i 38). En aquest període té lloc el creixement real del volum embrionari, principalment a partir de l'augment de volum del blastocele (Figs 39, 40, 41, 42, 43 i 44). L'embrió en aquest estadi s'anomena blastocist. En aquest estadi ja es distingeixen dos tipus cel·lulars, les cèl·lules del trofoectoderm i les cèl·lules de la massa cel·lular interna (Figs 45, 46, 47, 48 i 49). El blastocist posseeix una capa unicel·lular perifèrica de grans cèl·lules aplanades, el trofooctoderm o trofoblast (Figs 50, 51 i 52), i un grup de cèl·lules més petites en un punt de la cavitat central (Figs 53, 54 i 55). Aquest conjunt de cèl·lules s'anomena massa cel·lular interna i donarà lloc principalment a l'organisme adult, mentre que les cèl·lules del trofoectoderm formaran la placenta i les membranes embrionàries. El primer tipus cel·lular forma un veritable epiteli al voltant del blastocele, el que li dóna a l'embrió una aparença d'esfera buida, les parets de la qual, provocat per l'augment de volum progressiu del blastocele, es van aprimant mitjançant l'allargament de les cèl·lules del trofoblast (Fig 56). El segon tipus cel·lular, la massa cel·lular interna o embrioblast, es troba internament al trofoectoderm i únicament en un pol del blastocist (Figs 57 i 58).
No obstant, una característica dels embrions del marsupial és que no posseeixen massa cel·lular interna, desconeixent-se els factors que determinaran quines cèl·lules formaran l'embrió i quines formaran les membranes embrionàries.
Fins aquest estadi, els embrions de tots el mamífers s'assemblen molt. No obstant, els blastocists es desenvolupen de forma diferent en funció de grups d'espècies. Al ratolí, i probablement a l'home, el blastocele comença a formar-se quan l'embrió no te més de 20 o 30 cèl·lules (Figs 59 i 60), mentre que al moment en el qual la implantació comença, el blastocist conté no més de 100 cèl·lules (Fig 61). Al conill, la formació del blastocist comença a prop de tres divisions cel·lulars més tard que al ratolí, mentre que el blastocist expandit conté milers de cèl·lules i mesura de 3 a 4 mm en diàmetre (Figs 62 i 63). A l'ovella i la vaca, el blastocist s'allarga gradualment i pot arribar a una longitud de 20 cm abans de la implantació en la segona o tercera setmana de gestació (Fig 64 i 65). Als porcs, el procés d'elongació és més accentuat entre els dies 9 i 16 de gestació; el blastocist pateix un procés d'elongació 300 vegades més accentuat que en els remugadors, canviant d'una vesícula esfèrica petita a un tub allargat de més d'un metre de longitud, abans de que la veritable implantació comenci.
El moment en el qual la cavitat blastocèlica apareix no està relacionat amb la mida de l'embrió. Si 3 blastòmeres d'un embrió de 4 cèl·lules són destruïdes, la cèl·lula supervivent continua el seu desenvolupament, formant-se el blastocele en el moment usual, però donant lloc a un blastocist amb una talla 4 vegades inferior a un blastocist normal (Figs 66 i 67). Si un embrió és disgregat en cèl·lules aïllades, una vesícula plena de líquid apareix de forma sovint en les cèl·lules individuals en el moment en el que el blastocele es formaria, suggerint que el fluid s'origina per acumulació intracel·lular.
La característica més important del blastocist és la seva diferenciació en trofoectoderm i massa cel·lular interna. La diferenciació és el punt central del misteri encara no resolt del desenvolupament embrionari, ja que d'un sol òvul fecundat es formen tots els teixits d'un organisme adult, incloent diversos tipus com os, encèfal, espermatozous, etc. Les cèl·lules del trofoectorderm apareixen relativament especialitzades, són grans i aplanades amb nombrosos microvilli, formant la continua i tancada paret del blastocist. Aquestes cèl·lules constitueixen els 2/3 del total de cél·lules del blastocist. Investigacions amb microscopia electrònica han mostrat que les seves membranes estan íntimament unides i interdigitades, anclades en intervals per unions de tipus estret. Aquestes unions estretes ja han estat observades als embrions de 8 a 16 cèl·lules. Funcionalment, el trofoectoderm actua com una bomba, la qual és responsable del transport actiu d'ions entre el medi extern i el blastocist, i així és responsable indirecte del pas de fluid a l'interior del blastocele. Més tard, el trofoectoderm donarà lloc a cèl·lules trofoblàstiques primàries, les qual són les primeres en contactar amb el teixit matern, envaint les capes de l'endometri. En canvi, les cèl·lules de la massa cel·lular interna són petites, arrodonides, amb una alta tassa mitòtica, mostrant mínimes adhesions o especialitzacions fins que la implantació té lloc. La diferenciació també es veu manifesta a nivell bioquímic, ja que les cèl·lules de la massa cel·lular interna i el trofoectoderm sintetitzen diferents poblacions de proteïnes.
El següent teixit
a diferenciar-se és l'endoderm primari o hipoblast, el qual es forma a partir
de la capa de cèl·lules de la massa cel·lular interna
que està en contacte amb el blastocele. Les cèl·lules
del endoderm migren per tota la superfície interna del trofoectoderm.
Desprès de la implantació, aquestes cèl·lules
jugaran un important paper nutritiu per l'embrió, però no
donaran lloc al definitiu endoderm embrionari. La porció restant
de la massa cel·lular interna és ara anomenada epiblast,
el qual donarà lloc a la gastrulació a l'ectoderm, mesoderm
i definitiu endoderm embrionari.
5. Implantació: eclosió de la zona pel·lúcida
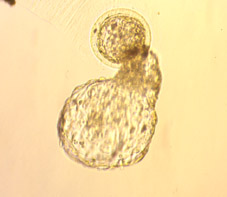
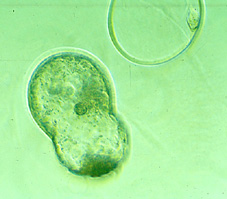
A l'inici de la implantació,
els blastocists de diverses espècies (no en el cas dels embrions
de conill) presenten l'eclosió espontània de la seva zona
pel·lúcida (Figs 68, 69,
70,
71,
72,
73,
74,
75 i 76). Aquest
és un procés d'eliminació d'una capa acel·lular externa
que ha mantingut en cohesió les cèl·lules durant el
desenvolupament preimplantacional i que probablement ha evitat la resposta
immunitària de l'úter matern.
En aquest moment s'inicia el verdader procés de implantació amb un canvi de tipus nutricional del embrió, que passarà de nutrir-se de les secrecions dels òrgans tubulars materns a dependre de la sang materna després de la placentación.
Aquesta obra finalitza amb el període d'implantació, fins aquest moment, l'embrió circulava lliurament pel tracte genital femení i permet la seva obtenció de forma no lesiva, fet que es utilitzat per a la aplicació de tècniques de reproducció assistida, incloent la transferència embrionaria.
Annex: Notes sobre el terme "embrió"
La reproducció
sexual dels animals mamífers es pot dividir de forma general en quatre grans períodes (Nomina Embryologica Veterinaria, N.E.V.,
1994). Aquests períodes són, cronològicament: la gametogènesi
i els períodes preembrionari, embrionari i fetal.
La gametogènesi
inclou els processos d'espermatogènesi i ovogènesi.
El període preembrionari engloba una sèrie de processos que van des de l'ovulació i inseminació o còpula fins a la neurulació, passant per la fecundació, la blastogènesi o segmentació, la blastulació i la gastrulació.
Posteriorment, la formació del tub neural porta l'inici del període embrionari, en el qual es fa més correcte la utilització del terme "embrió". No obstant, l'expressió "embrió" s'utilitza en la majoria d'ocasions com a sinònim d'aquella estructura reproductiva ja fecundada que es manté en relació amb el tracte reproductor genital femení, tant immersa en les seves secrecions en els estadis inicials com després de la implantació.
En ocasions, el terme embrió sol ser qualificat com a preimplantacional o en fase de preimplantació per definir que aquest es troba lliure en les secrecions oviductal o uterina. D'una forma més concreta, el terme embrió se sol acompanyar del nombre de cèl·lules que el composen en aquell instant (és a dir, un embrió compost per quatre cèl·lules se sol denominar embrió de quatre cèl·lules o en estadi de quatre cèl·lules), des de que es troba com una cèl·lula única (embrió d'una cèl·lula o zigot) fins que posseeix de 16 a 32 cèl·lules. A partir d'aquest moment el terme embrió se sol substituir pel de mòrula o blastocist, segons si es troben un nombre de cèl·lules compactades que no permet un recompte senzill o s'ha format una cavitat central plena de líquid anomenada blastocele, respectivament.
Diversos autors han intentat estendre l'ús d'un terme més correcte que el d'embrió per definir aquesta estructura preembrionària en fase preimplantacional. D'aquesta manera, Whittingham (1975) va emprar l'expressió d'embrió jove ("early embryo"). Per la seva part, Jones (1990), motivat per la dificultat d'establir els límits ètics i morals de la manipulació d'embrions humans, va optar per utilitzar el terme de preembrió ("pre-embryo"). Jones va definir aquesta estructura com aquella existent des del procés de fecundació fins a l'aparició de la línia primitiva, quan ja ha tingut lloc la implantació. No obstant, la formació dels solcs i plecs neurals, de la notocorda, dels somits, de les cúpules òptiques i de la placoda òtica tenen lloc també sota el període embrionari (N.E.V., 1994). Barone (1978), en el seu estudi descriptiu del desenvolupament de la gestació, va utilitzar els termes "d'ou segmentat", mòrula i blastocist per definir les estructures de la fase de preimplantació. Aquesta terminologia va ser la més emprada en els estudis embriològics de l'època. No obstant, Betteridge (1977) va utilitzar el terme embrió com sinònim tant d'ovòcit fecundat com de mòrula, de blastocist o de blastocist expandit. Aquesta última accepció del terme embrió és la que ha tingut una major utilització en els estudis experimentals reproductius i de reproducció assistida, tal com el cultiu in vitro i la crioconservació d'embrions.
El terme utilitzat
en el present estudi per definir aquella estructura reproductiva, ja fecundada
i en fase de preimplantació (ja sigui amb localització oviductal
o uterina), ha estat el d'embrió i qualificat com a preimplantacional
o concretant el seu estadi. L'ús d'aquest terme es veu motivat per
la seva gran extensió en la terminologia reproductiva, el que comporta
que aquest ús tingui com objectiu una major coherència i
sincronia amb la terminologia majoritàriament emprada.
{kind=link}